

INTRODUCTION
Fecal incontinence (FI) can be a devasting and stigmatizing disease for elderly patients [1-3]. Age is associated with deteriorated anorectal functions presenting with lower anal resting and squeeze pressures, anal sphincter denervation, increased rectal compliance and decreased rectal sensation [4-6]. In addition, dementia, mobility impairment, comorbid chronic diseases, and bowel disturbances including diarrhea and constipation are the most common independent risk factors for FI [7]. Nontraumatic internal anal sphincter (IAS) weakness observed frequently in elderly FI patients is different in pathophysiology from patients with external anal sphincter (EAS) injuries during delivery [8, 9] or iatrogenic injuries during perianal fistula or anal fissure surgery [10, 11]. Unlike in younger FI patients, in elderly FI, age-related phenotypes are characterized by lower resting pressure, however, 2 have similar squeeze pressure [12].
It is also known that resting pressure maintained by the basal tone of IAS is a main clinical parameter of FI [13, 14]. Compared to the voluntary contractility of skeletal structures of EAS, IAS, conjoined at the end of the gastrointestinal (GI) tract, has autonomic nerve fibers distinct from somatic fibers entering the EAS via the pelvic plexus [15]. Therefore, as an intrinsic property, the smooth muscle (SM) of IAS plays a critical role in maintaining basal tone independent of extrinsic nerve stimulation. Various studies related to restoration of SM cells (SMCs) of IAS as FI treatment have been evolved. In vitro and in vivo experimental studies using bioengineered human IAS have reported that implantation of intrinsically innervated human IAS constructs generated spontaneous basal tone with myogenic and neuronal components [13, 14]. Furthermore, FI cell treatments [16-19] using bone mesenchymal stem cells (MSCs), adipose stem cells (ASCs), myoblasts, and a combined treatment strategy [20-24] have been reported functional improvement in the injured sphincter. However, these studies for cell therapy showed a bias and lacked data for clinical use as reported by the previous systematic reviews [25-27]. Data on the regeneration of IAS associated with basal tone was also limited, and most cell therapy studies were performed on animal models for the skeletal muscle of EAS or nerve damage [25, 27, 28].
As an intrinsic force of the sphincter SM, the basal tone is generated from the cross-bridge movement of myosin and actin filaments on 20-kDa myosin regulatory light chain phosphorylation (p-MLC20) [29]. Several studies have demonstrated that RhoA and Rho-associated, coiled-coil containing serine/threonine kinase (ROCK) is the major molecular determinant of basal tone in IAS [30, 31]. Recently, it has been covered that multifactorial pathways are involved in the modulation of basal tone including calcium transients or aging-associated IAS dysfunction (AAID) [32, 33]. Some authors have suggested a need for an integrative approach for the uncovered molecular mechanism with a new genesis of spontaneous basal tone in IAS [34]. Furthermore, the IAS relaxation, which is critical for the rectoanal inhibitory reflex (RAIR) in response to defecation, is highly dependent on nerve activity with inhibitory neurotransmitters (NTMs) compared to the ability of IAS to develop spontaneous tone, independently [32, 33].
However, the complexity in the molecular mechanism of myogenic features in IAS might limit preclinical or clinical outcomes of FI cell therapy or regenerative medicine. These treatment modalities should entail the molecular basis of SMCs, which generate basal tone as an intrinsic property and the relaxation in IAS [31]. So far, there are no systematic reviews for translation studies on IAS-based FI treatment covering molecular pathways, regenerative medicine, and cell therapy. This study aims to perform a systematic review of the literature for a comprehensive understanding of the IAS as a critical role in incontinence in FI patients and provide the interrelationship of those studies.
METHODS
Literature search
To identify all the available translation and experimental studies on the IAS in FI published and indexed up to January 31, 2022, a systematic search was conducted in PubMed, Cochrane Library, EMBASE, Medline, and Web of Science. This strategy included in vitro and in vivo (animal or human) translation studies related to cell therapy, regenerative medicine, tissue engineering, and preclinical study. Medical Subject Headings (MeSH) terms and Emtree terms were used in PubMed and EMBASE, respectively, together with separate words or word combinations in the title or abstract.
Type of article reviewed and data extraction
Automatic deduplication of the retrieved articles was performed twice with EndNote (ver. X9.0.1, Clarivate Analytics, Philadelphia, PA, USA); first per the searched database, and then based on the relevant titles with the assessment of IAS SMCs-related terms including the basal tone, the relaxation, cell therapy, and regeneration medicine. Studies including anatomy and histology were excluded. Two independent reviewers extracted the data from the designed extraction form. Translation studies for SMCs of IAS were classified using keywords as follows: calcium, Rho/ROCK, AAID, oxidative stress, renin-angiotensin-aldosterone, cyclooxygenase (COX), and inhibitory NTM (Table 1). The following data were collected: tissue sample for harvest, mediator, pathway, the action of mechanism in translation studies; cell type, animal model, type of sphincter injury, confirmation of incontinence, implantation, scaffolding, cell tracking, and outcomes measurements in the experimental studies for IAS treatment.
RESULTS
Intrinsic molecular pathway of basal tone in internal anal sphincter
Calcium
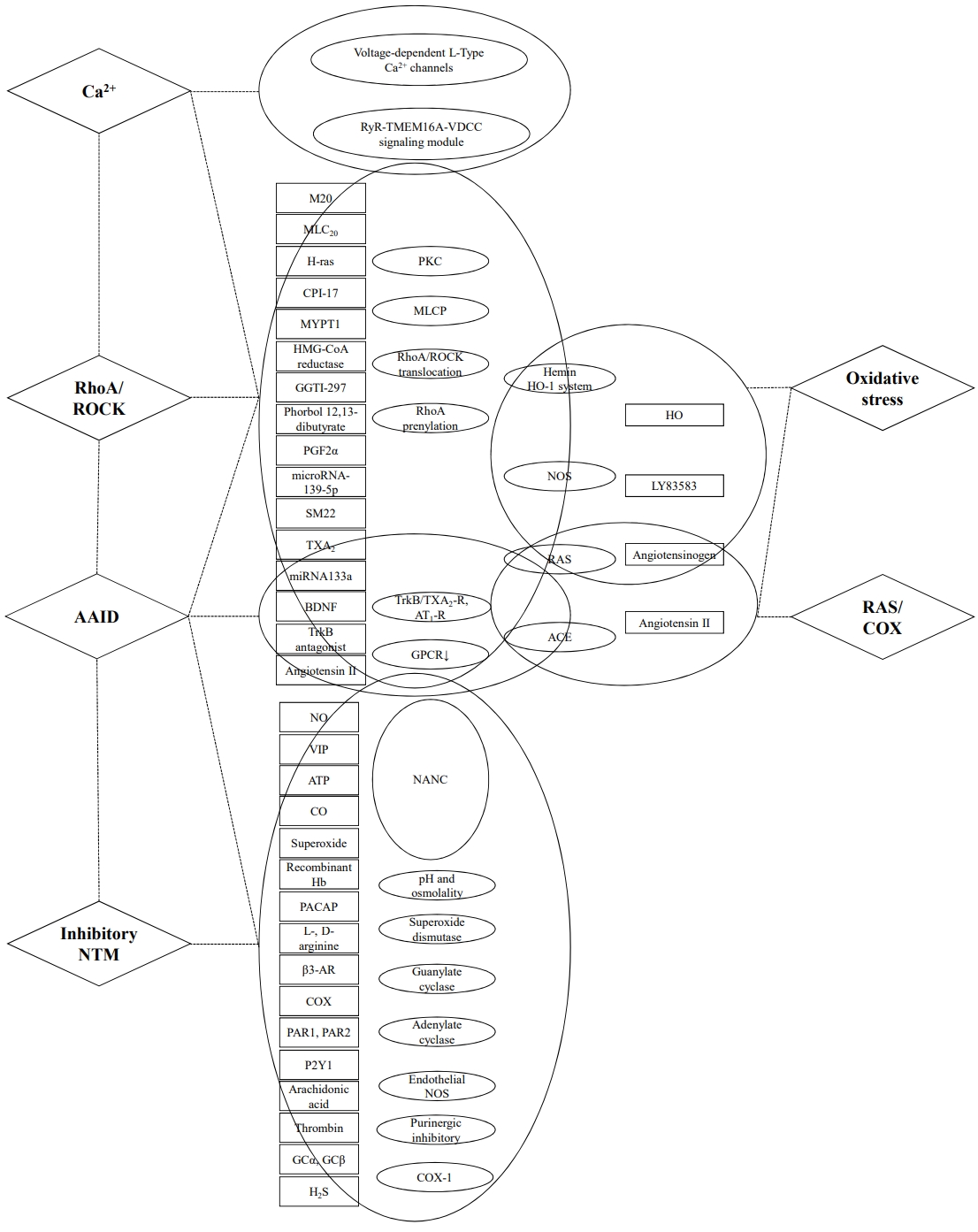
Calcium (Ca2+) activation in Ca2+/calmodulin/myosin light chain kinase (MLCK) pathway plays a critical role in the initial phasic stage of IAS tone development via G protein-coupled receptor (GPCR) activation, and in promoting the p-MLC20 [31]. Ca2+-related factors (endothelin, Ca2+-free solutions and Ca2+-channel blockers, Ca2+ oscillations, Ca2+ transients [CTs], and Ca2+-mediated signaling cascade) can determine the myogenic intrinsic property of the basal tone in IAS directly, rather than extrinsic nerve and hormone stimulation. The intracellular mechanism of endothelin is the biphasic action characterized by an initial brief relaxation followed by a sustained contraction via inhibition of selective protein kinase C (PKC) inhibitor H-7 or calmodulin inhibitor W-13 [35]. Furthermore, a recent study reported an impaired global rise of Ca2+ in animal models using mice with knockout SM-specific myosin phosphatase target subunit 1 (MYPT1), which anchors myosin light chain phosphatase (MLCP) to p-MLC. The study demonstrated that maintaining IAS basal tone requires Ca2+-mediated MLCK activation [34]. Three channels are involved in generating the Ca2+ rise; Ca2+-releasing ryanodine receptors/channels (RyRs), transmembrane member 16A (TMEM16A) Ca2+-activated Cl (ClCa) channels, and L-type voltage-dependent Ca2+channels (VDCCs). Therefore, altering any one of these channels impaired IAS basal tone similar to the MLCK deletion. This suggests the independence of molecular mechanisms for the initial phase of IAS tone development from the later phase of maintenance of the basal tone. The study also showed the potential of a new treatment targeting a global rise in Ca2+-mediated signaling cascade (RyR-TMEM16A ClCa channel-L-type VDCC) [34].
Serving as the pacemaker, Ca2+-activated ClŌłÆ channel TMEM16A also propagates electrical slow waves (SWs) into IASŌĆōSMCs, inducing basal tone via the depolarization and activation of L-type VDCCs and giving rise to CTs in the SMCs. The pharmacological and spatiotemporal properties of CTs in the IAS are very similar to those of SWs with the greatest frequency and amplitude at the distal end of the IAS, suggesting that this is the predominant pacemaker region. Phasic activity by the fastest and highly coordinated pacemaker located at the distal extremity will lead to orally directed contractions that will aid in maintaining continence [36]. The basal tone generated by RyR-TMEM16A Cl Ca2+ channel-L-type VDCC signaling module might also be mediated by oscillating Ca2+ signals in SMCs of IAS [32]. The opening of a small number of RyRs induced Ca2+ release with an exhibition of asynchronized Ca2+ oscillations (ACaOs) and synchronized Ca2+ oscillations (SCaOs) in IAS. In sustained contractions, ACaOs contribute approximately 25%, while SCaOs account for 75%.
Rho/Rho-associated, coiled-coil containing serine/threonine kinase pathway
The basal tone in the IAS is critically dependent on RhoA/ROCK [37]. The balance between the Ca2+/calmodulin/MLCK stimulation and MLCP inhibition determined primarily by RhoA/ROCK pathway plays a crucial role in the latter phase of tonic followed by the initial phasic stage of IAS tone development [31, 38]. The latter phase of tonic or sustained contraction in basal tone is dependent on the MLCP inhibition which maintains higher levels of p-MLC20 [39]. RhoA/ROCK which is present throughout the SMCs translocates to the periphery of the SMCs during the sustained contraction phase.
It has been reported that several factors are involved in the regulation of basal tone in IAS via modulating the molecular pathway of RhoA/ROCK in the previous studies [38-45]. The spontaneous tone in the IAS can be induced by upregulation of RhoA/ROCK and PKC in part which mediates its effects via ROCK [38, 39]. Singh et al. [43] reported that phorbol esters, which have been used as PKC activators in SMs, cause contraction of the human IAS SMCs associated with RhoA/ROCK activation rather than the PKC activation. Furthermore, RhoA prenylation is essential for the translocation of RhoA to the cellular membrane and downstream signal transduction of RhoA. Consequently, inhibition of RhoA prenylation by geranylgeranyltransferase I inhibitor, inactivates RhoA/ROCK, and it leads to a decrease in the basal tone in the IAS [41]. Rattan [42] reported that the HMG-CoA reductase inhibitors such as simvastatins relax the IAS SM via decreasing the levels of RhoA prenylation and ROCK activity. de Godoy et al. [40] also reported that H-ras modulates basal tone by inhibiting RhoA translocation to the plasma membrane, reducing activation of the Rho kinase isoform ROCK II. Inhibition of upstream RhoA-Rho kinase signaling produces downstream prerelaxation events including diminished p-MYPT1 and increased activity of MLCP, resulting in diminished p-MLC20. Thromboxane A2 (TXA2), prostaglandin F2╬▒ (PGF2╬▒), and angiotensin II (ANG II), which are the end products of arachidonic acid and renin-angiotensin system (RAS) pathway, provide extracellular signals that trigger the activation of RhoA/ROCK for the basal IAS tone in humans [44]. The microRNA (miRNA)-139-5p plays a significant role in the functional phenotypic SMs by repressing RhoA/ROCK pathway in the SMCs, and its low expression in the IAS may increase the basal tone [45]. Phosphorylation of SM22 (transgelin/calponin) protein by ROCK inhibiting SM22-actin interaction leads to the development of basal tone in IAS [46]. The selective ROCK inhibitor which causes relaxation of the IAS might be used for conditions associated with hypertensive IAS [37].
Aging-associated internal anal sphincter dysfunction
Clinically, degenerative transformation of the anal sphincter muscles can be characterized by decreasing IAS tone and increasing response to RAIR and subsequent decreased resting pressure rather than squeezing pressure. Recently, some authors defined AAID with the molecular mechanisms and impeded the development of a specific and safe treatment for AADI-related FI [33, 47]. A previous study reported that aging can induce the downregulation of GPCR via TXA2 and ANG II type 1 receptors desensitization (TXA2-R and AT1-R). In their data, the fibroelastic properties (FEP) of the SMs correlated with a decrease in the GPCR expression at gene and protein levels, and an increase in the expression of GPCR downstream signaling regulatory proteins, RGS2, GRK5, and Arrb2. Therefore, downregulation and compromise in the GPCR recycling lead to a decrease in the IAS tone [48]. An endogenous agent such as a brain-derived neurotrophic factor (BDNF) may also relate to the AAID. As an important member of the neurotrophin family, BDNF which is released primarily in the central nervous system was also released in the peripheral nervous system, especially in the enteric nervous system and in the GI tract SMs. Using different systems, BDNF can rescue AAID via augmentation of GPCR-coupled SM contractility and relaxation. BDNF-mediated increased basal tone can be induced in GPCR-coupled agonist-stimulated IAS by activation of RhoA/ROCK in the SMCs, and via a decrease in the nitrergic nonadrenergic noncholinergic (NANC) inhibitory neurotransmission [33]. BDNF also increased the basal and agonist-stimulated IAS tone via TrkB/TXA2-R and TrkB/AT1-R signaling, and RAIR-induced IAS relaxation via nitric oxide (NO)/soluble guanylate cyclase (GC) pathway [47].
Furthermore, the miRNA-133a and its gene targets are crucial to the RhoA signaling pathway in relation to contractility and IAS SM phenotype in aging [49]. In their data, it was observed that the basal tone and the FEP of the IAS might be regulated by changes in the transduction molecules, growth and transcription factors, regulators of SMC differentiation, extracellular matrix components, and early markers of SMCs via miRNA-targeting diverse signaling pathways [49]. Most of the AAID studies have been performed on animal models using older rats or compared with younger ones as our review, showing an impossible approach for harvesting sphincter tissue from younger and elderly FI patients.
Oxidative stress
The antioxidant defense mechanisms, such as superoxide dismutase (SOD), catalase, and glutathione peroxidase play a crucial counterregulatory role in ROS levels which are produced during aerobic cellular metabolism. However, with aging, the weakening of the antioxidant defense mechanism can bring about cumulative effects of excessive ROS and results in irreversible cellular damage. As an aging-related risk factor, oxidative stress has been reported to decrease the overall propulsive activity of the gut. A decrease in IAS tone and altered gut motility may be the leading causes of AAID via decreasing phosphorylation of the myosin-binding subunit of MYPT1 and regulatory MLC (MLC20), secondary to RhoA-associated kinase (RhoA/ROCK) inactivation [50-52].
Singh et al. [50, 52] also reported that the effect of extreme oxidative stress decreases IAS tone, via inactivation of RhoA/ROCK II in rats, while a lower level of oxidative stress leads to an increase in IAS tone. Antioxidants such as SOD might play an important role in reversing the aging-associated oxidative damage in IAS. Furthermore, heme oxygenase (HO) degrades heme, producing carbon monoxide, biliverdin, and Fe2+ [43]. Krishna et al. [51] identified the role of HO in neurally mediated relaxation of IAS. It was shown that carbon monoxide (CO), one of the HO products, causes SM relaxation along with hyperpolarization of the SMCs via upregulation of RhoA/ROCKII and downregulation of MYPT1 levels.
Molecular pathway of relaxation in internal anal sphincter
Renin-angiotensin system and cyclooxygenase
In the pathway of the RAS, renin converts angiotensinogen into angiotensin I, and angiotensin I is converted into ANG II by the angiotensin-converting enzyme (ACE). ANG II activates specific membrane receptors in the target cells, and binds to 2 different subtypes of GPCRs; AT1-R and AT2-R. Previous studies demonstrated that RAS via ANG II biosynthesis inhibitor (ACE inhibitor) and release of ANG II followed by activation of AT1-R and AT2-R, provide a partial regulation of the basal tone in the IAS [53, 54]. ANG II can also produce a bimodal effect, a contraction (at lower concentrations) followed by relaxation (at higher concentrations) [55]. A contraction of IAS is mediated by AT1-R, and relaxation of IAS is mediated by AT2-R. Higher concentrations of ANG II result in the internalization of the AT1-R to the cytosol and the externalization of AT2-R to the plasm membrane. The translocated AT2-Rs for activation exert their inhibitory effect on the SMCs of IAS leading to relaxation. In addition, among the 2 primary isoforms of the COX pathway, COX-1 might play a role in maintaining basal tone by producing PGF2╬▒, thromboxanes, and ANG II which provides the signal at SMCs of IAS [56, 57]. However, the molecular mechanism between RhoA/ROCK and PKC in the COX-1ŌĆōsustained tone in the IAS remains unclear.
Inhibitory neurotransmitter
IAS relaxation presented as RAIR in response to defecation is dependent on nerve activity with NTMs [58]. In functional experiments, noradrenaline is the only NTM that has been identified to elicit an excitatory response in the sphincter, while inhibitory roles have been suggested for NO, CO, vasoactive intestinal polypeptide (VIP), acetylcholine and adenosine 5ŌĆÖ-triphosphate (ATP), and pituitary adenylate cyclase-activating peptide (PACAP) [59-64]. These transmitters appear to interact for SM relaxations, especially NO. NO which is synthesized from L-arginine in a reaction catalyzed by NO synthase (NOS), is freely soluble, diffuses rapidly, has a short half-life (3 seconds) [65, 66], and mediates neurogenic relaxation by binding to cytosolic GC and stimulating the production of cyclic guanosine monophosphate (cGMP) [67]. In absence of NOS , other inhibitory NTM including VIP or ATP, heme compensates to maintain an approximately normal response [68]. NO also caused the IAS relaxation after activation by the formation of NO3ŌĆō after contact with the superoxide [69] or via proteinase-activated receptor-1 (PAR1) and PAR2 [70].
The interstitial cells of Cajal (ICC), which are located in close proximity to the nerves throughout most of the GI tract including IAS, participate in nitrergic and cholinergic NTM in the IAS. In other interstitial cells, platelet-derived growth factor ╬▒ (PDGFR╬▒) is thought to be responsible for the purinergic component of inhibitory NTM. ICC and PDGFR╬▒ cells are both closely associated with neural NOS neurons in the IAS. SMCs from gap junctions with adjacent ICC and PDGFR╬▒ cells, give rise to an electrical syncytium referred to as the ŌĆ£SIP syncytiumŌĆØ. The nitrergic relaxation in the IAS results from the signals integration by GC within the SIP syncytium, while cGMP-dependent protein kinase I and its downstream targets are responsible for less than half of the relaxation [71]. In addition, endothelial NOS may partly modulate the vascular SMs relaxation in response to activation of the ╬▓3-adrenoceptor (AR). The stimulation of ╬▓-AR causes a release of Gs╬▒, which activates adenylate cyclase to produce cAMP. Elevated levels of cAMP produce SM relaxation in IAS via activation of cAMP-dependent protein kinase A [72]. A NO-like substance (CO, and VIP) as an inhibitory NTM of NANC nerves, is responsible for the neutrally mediated IAS SM relaxation [67, 73, 74]. Rattan et al. [59] compared the ability of NO, CO, and VIP to relax mouse IAS. VIP, which mediated ultraslow relaxation and hyperpolarization by purinergic and nitrergic NTM, was the most potent, followed by NO. While, longer stimulus trains of electrical field stimulation, a non-purinergic, non-nitrergic relaxation and hyperpolarization, which is mediated by VIP, lead to prolonged IAS relaxation [75], and very high concentrations of CO may not be a relevant relaxation agonist in mouse IAS [59]. The purines such as ATP also contributed to inhibitory NTM in the IAS. Duffy et al. [76] reported that purinergic hyperpolarization associated with IAS relaxation is independent of intramuscular ICC. McDonnell et al. [60] reported that a purinergic pathway significantly contributes to inhibitory motor innervation in the IAS by hyperpolarization of the postjunctional membrane to produce inhibitory junction potentials. Folasire et al. [61] also reported that hydrogen sulfide possibly contributes relaxation of the porcine IAS, and its potency is lower than that of CO. In addition, recombinant hemoglobin 1.1 (rHb1.1) could be caused specific obliteration of NO-induced relaxation of the IAS [77]. However, nitrate tolerance did not develop in IAS SM in the previous study [78].
Internal anal sphincter treatment in fecal incontinence: cellular vs. acellular
Inoue et al. [16] reported that cell treatment using ASC sheets in an animal model showed a significant improvement of the anal pressure in the ASC sheet transplantation group compared with the control group after 8 weeks of cell implantation. Histologically, in the vicinity of the remaining SM, reproduction of SM was detected with an increased level of SMC-specific markers. Cell treatment using human ASCs (hASCs) injections combining them with polyacrylamide hydrogel carrier, bulkamid showed the functional improvement in anorectal manometry, low inflammatory response, and tissue integration on micro-computed tomography [19]. Salcedo et al. [17, 18] also reported a significant increase in resting anal sphincter pressure and peak pressure, anal sphincter electromyography amplitude and frequency, and a marked decrease in fibrosis and scar tissue in an animal model using an intramuscular or intravascular injection of MSCs. Oh et al. [20] showed the functional differentiation of polycaprolactone beads containing autologous myoblasts labeled with PKH-26 fluorescent dye in animal models using posterior resection of IAS and EAS. But it remained weak and unclear for an association between functional improvement of resting pressure and the differentiation of myoblasts into SMCs of IAS. Sarveazad et al. [79] reported the efficacy of co-application of laser and hASC therapy showing improved resting anal pressure and enhanced myogenesis and angiogenesis with increased expression level. All the previous incontinence models included incision of EAS or adding pudendal nerve crush [16-20, 79]. In the observed histologic augmentation of smooth muscular layers in IAS, but in cell tracking, no newly differentiated SMCs of implanted stem cells were found at the injury site under fluorescence in situ [16], micro-computed tomography [19], confocal microscopy, or immunofluorescence assay using green fluorescent protein labeling [17, 18] (Table 2).
In contrast to cellular treatment, scaffolding or gut engineering using natural biomaterial has been employed for IAS malfunctions [13, 14, 80-83]. Some authors developed a 3-dimensional (3D) bioengineered ring (BR) model constructed from isolated human IAS SMC [80-82] (Table 2). Bioengineering an in vitro 3D fibrin-based model consisting of a homogenous population isolated from human sphincteric SMC on a fibrin gel might provide a favorable cellular environment to cell survive tissue expansion compared to the previous cell treatment [16, 18-20, 22, 69]. The 3DBR showed an increase in contractile protein expression profiles of PKC, RhoA, CPI-17, and HSP27 which have the specificity of basal tone generated by IAS circular SM at rest [81]. In the IAS reconstructs, the basal tone was maintained with developing spontaneous tone (0.68 ┬▒ 0.26 mN), and concentration-dependent tone response to bethanechol (a muscarinic agonist), K+ depolarization, and isoproterenol (╬▓-adrenoceptor agonist), Y-27632. It also had higher levels of RhoA/ROCK, PKC-potentiated inhibitor, or inhibitory phosphoprotein for myosin phosphatase (CPI-17), MYPT1, and MLC20 comparted to rectal SM [84]. In intrinsically innervated human IAS models using enteric neuronal progenitor cells isolated from the human colorectum, also demonstrated that myogenic and neuronal components of the bioengineered IAS response physiologically to VIP-ergic, electrical stimulation, cholinergic, KCl-induced neurotransmission with relaxation, and contraction of basal tone [13]. However, it is limited due to complications such as infection and erosion related to foreign body reaction at perineal implantation [14], and no a clinical trial on bioengineered IAS reconstructs has been reported.
The interrelationship between the molecular mechanism and regenerative treatment for internal anal sphincter
Several translation studies have revealed the molecular mechanism of SMC in IAS as intrinsic properties, which can be characterized by Ca2+, RhoA/ROCK, AAID, oxidative stress, RAS, COX, and inhibitory NTM. However, we observed that previous cellular treatments [16-20, 79] did not show a relationship between SMCs-related histologic or functional outcomes and basal tone or relaxation-related molecular pathways in injured IAS regardless of the cell type or confirmation of IAS-incontinence. We considered that it might be due to the complexity of the molecular mechanisms for basal tone and relaxation of IAS as intrinsic properties (Fig. 1). While the bioengineered IAS reconstructs as acellular approach models have a potential for in vivo response and functional properties of 3DBR similar to human IAS dependence on RhoA/ROCK [84].
The best option of cell source and phenotype is still a questionable issue in the field of GI regeneration [85]. The myogenic features SMCs isolated from IAS are different from those of skeletal muscle cells from EAS in the human anal sphincter [86]. Therefore, cell treatments using cell types originated from skeletal muscle might be unsuitable for restoration of dysfunctional IAS clinically, which is composed of SMCs characterized by intrinsic force contracted at rest, with small-amplitude slow contractions and slow relaxation. These implied that only one study using autologous skeletal-muscle-derived cells showed a significant improvement in resting pressure after cell implantation, but it did not demonstrate the physiologic relationship between those outcomes and basal tone of IAS as a role to maintain resting pressure [87]. However, according to a recent study, myogenic progenitor cells derived from the skeletal muscle can be differentiated into SMCs in vitro culture of transforming growth factor ╬▓1 and heparin-containing medium, with showing contractile proteins expression profiles and functional voltage-gated ion channels associated with SM lineage commitment [88].
Moreover, in the included studies, experimental evidences performed on real human IAS tissue were only 5, and most animal models used rats or mice with contractile protein-specific gene deletion or inhibitory drug. Harvest of viable SMCs of IAS isolated human anal sphincter might be quite difficult because only patients who underwent abdominal perineal resection operation can provide intact anal sphincter. A larger-scale human anal sphincter also needs a new approach for expansion capacity, for example, 3D scaffolding or spheroid culture, and organoids, reflecting that the previous studies have been performed on small size animals like rats or mice. In limitation, we could not performe an interrelationship analysis based on experimental methods for between included molecular mechanisms and regenerative treatment for IAS. Futher study needs with an interrelationship analysis for those.
CONCLUSIONS
This study implicates the complexity of the molecular mechanisms for basal tone and relaxation of IAS as intrinsic properties, which change from normal continence into incontinence in FI patients influenced by multifactorial etiology. Many translation studies have revealed the key pathway of IAS for the functional improvement of resting pressure and RAIR in FI. However, experimental evidence of FI cellular treatment has lower specificity for IAS, and a lower interrelationship between the molecular mechanisms of IAS and functional outcomes of cell treatments. Furthermore, there are no reports on lab-grown engineered gut tissues in patients with FI. The field of IAS regeneration presents more challenges and warrants more advances.